HOME | TEAM | RESEARCH | TEACHING | PUBLICATIONS
Research Focus
Plants adapt to their biotic and abiotic environment with numerous physiological and molecular responses. Our research group is interested in the metabolic and biochemical events that regulate plant-environment interactions. In particular, we investigate interactions of plants with pathogenic microorganisms in context with a plant immune response designated as systemic acquired resistance (SAR). SAR is induced by a localized leaf-inoculation with a pathogen and confers disease resistance against a broad-spectrum of phytopathogens systemically in the foliage. After SAR induction, plants are able to activate defense responses more quickly and vigorously when subsequently attacked (defense priming) (Figure 1).

- Prof. Dr. Jürgen Zeier
- Head of Institute
- +49 211 81-14733
- Molekulare Ökophysiologie
der Pflanzen
Heinrich-Heine-Universität
Düsseldorf
Universitätsstraße 1 - Building: 26.14
- Floor/Room: 02.075
- Eva Wenndorf
- Secretary
- +49 211 81-13505
- Building: 26.14
- Floor/Room: 02.067
Over the last decade or so, our work has significantly contributed to the formation of a consistent picture on the critical molecular events that underlie SAR and has clarified several controversies in the field (e.g. Mishina & Zeier, 2006; Mishina & Zeier, 2007a; Attaran et al. 2009; Návarová et al. 2012; Bernsdorff et al., 2016; Gruner et al., 2018; Hartmann et al., 2018). This has been achieved by the combinatory use of interdisciplinary methods such as physiological, genetic, biochemical, analytical-chemical, spectroscopic, and high-end metabolite profiling analyses. The spectrum of plant metabolites that we have analytically and functionally investigated in context with plant-biotic interactions and SAR is diverse and includes phenolics such as salicylic acid (SA) and its derivatives (Attaran et al., 2009; Bernsdorff et al., 2016), indolics (Stahl et al., 2016), terpenoids (Attaran et al., 2008), oxylipins (Mishina & Zeier, 2007a), sterols (Griebel & Zeier, 2010), tocopherols (Stahl et al., 2019), proteinogenic and non-protein amino acids (Návarová et al. 2012; Bernsdorff et al., 2016), and amino acid derivatives (Vogel-Adghough et al., 2013; Hartmann et al., 2017; Hartmann et al., 2018; Stahl et al., 2019) (Figure 2).
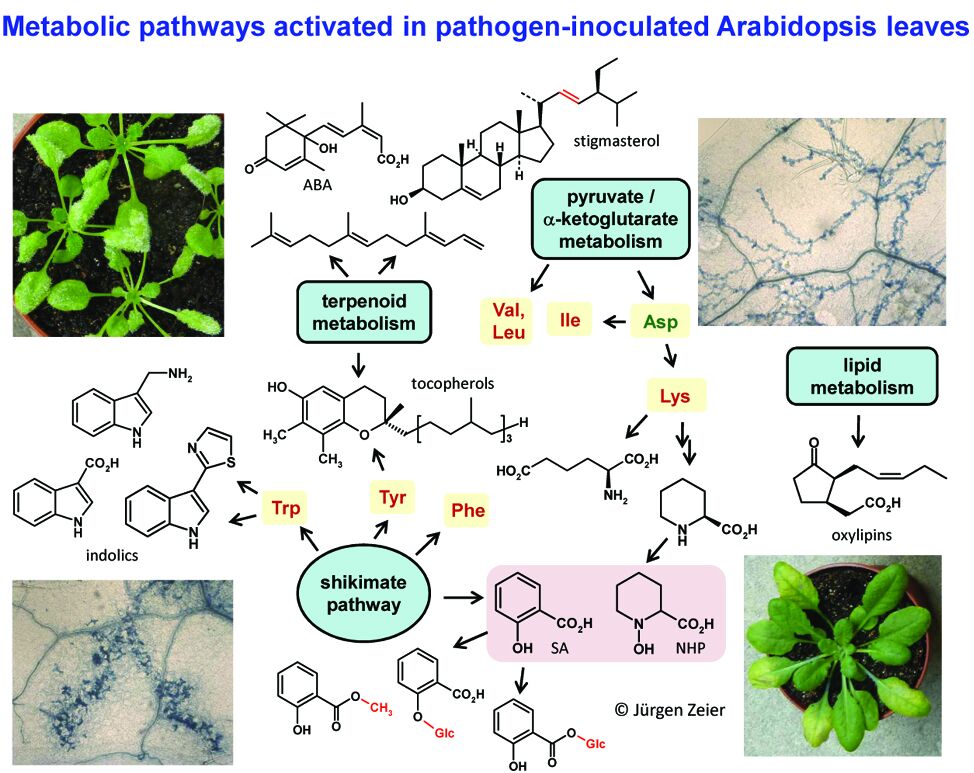
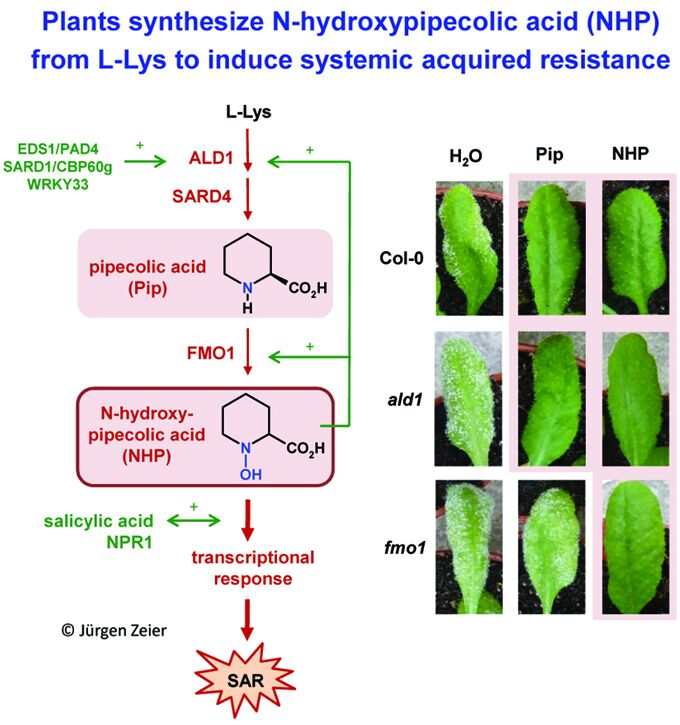
Significantly, we have identified a pathogen-inducible L-Lys catabolic pathway that generates N-hydroxypipecolic acid (NHP) as a critical endogenous regulator of SAR and the associated defense priming phenomenon (Hartmann & Zeier, 2018; 2019). NHP is biosynthetically derived from the non-protein amino acid pipecolic acid (Pip) (Návarová et al. 2012; Zeier, 2013; Bernsdorff et al., 2016), a lysine catabolite (Figure 3). We have biochemically clarified a three-step biosynthetic pathway by which NHP is formed from L‑Lys (Hartmann et al., 2017; Hartmann et al., 2018). The Arabidopsis aminotransferase AGD2-LIKE DEFENSE RESPONSE PROTEIN1 (ALD1) first abstracts the a-amino group of L-Lys to generate cyclic 2,3-dehydropipecolic acid (2,3-DP). 2,3-DP is then reduced to Pip by the NAD(P)H-dependent reductase SARD4/ORNCD1 (Hartmann et al., 2017). In a third step, the monooxygenase FLAVIN-DEPENDENT-MONOOXYGENASE1 (FMO1), which we had been more previously identified as a critical component of SAR (Mishina and Zeier, 2006), catalyzes the N-hydroxylation of Pip to NHP (Hartmann et al., 2018). As the end product of this “pipecolate pathway”, NHP acts as the actual SAR-inducing metabolite that primes plants for enhanced defense activation (Figure 3). The mode of action of NHP also involves direct induction of SAR gene expression, signal amplification, and positive interplay with salicylic acid signaling to ensure elevated plant immunity and SAR (Hartmann et al., 2018; Hartmann & Zeier, 2018; 2019). Since exogenous application of low doses of NHP provides strong pathogen resistance, our research might result in the development novel, natural-product-based plant protection strategies (link news) (Figure 3).